THE EVOLUTION OF MAN
Volume II
CHAPTER XVII
EMBRYOLOGY OF THE LANCELET AND THE SEA-SQUIRT
The structural features that distinguish the
vertebrates from the invertebrates are so prominent that there was
the greatest difficulty in the earlier stages of classification in
determining the affinity of these two great groups. When scientists
began to speak of the affinity of the various animal groups in more
than a figurative—in a genealogical—sense, this
question came at once to the front, and seemed to constitute one of
the chief obstacles to the carrying-out of the evolutionary theory.
Even earlier, when they had studied the relations of the chief
groups, without any idea of real genealogical connection, they
believed they had found here and there among the invertebrates
points of contact with the vertebrates: some of the worms,
especially, seemed to approach the vertebrates in structure, such
as the marine arrow-worm (Sagitta). But on closer study the
analogies proved untenable. When Darwin gave an impulse to the
construction of a real stem-history of the animal kingdom by his
reform of the theory of evolution, the solution of this problem was
found to be particularly difficult. When I made the first attempt
in my General Morphology (1866) to work out the theory and
apply it to classification, I found no problem of phylogeny that
gave me so much trouble as the linking of the vertebrates with the
invertebrates.
But just at this time the true link was discovered, and at a
point where it was least expected. Towards the end of 1866 two
works of the Russian zoologist, Kowalevsky, who had lived for some
time at Naples, and studied the embryology of the lower animals,
were issued in the publications of the St. Petersburg Academy. A
fortunate accident had directed the attention of this able observer
almost simultaneously to the embryology of the lowest vertebrate,
the Amphioxus, and that of an invertebrate, the close affinity of
which to the Amphioxus had been least suspected, the Ascidia. To
the extreme astonishment of all zoologists who were interested in
this important question, there turned out to be the utmost
resemblance in structure from the commencement of development
between these two very different animals—the lowest
vertebrate and the mis-shaped, sessile invertebrate. With this
undeniable identity of ontogenesis, which can be demonstrated to an
astounding extent, we had, in virtue of the biogenetic law,
discovered the long-sought genealogical link, and definitely
identified the invertebrate group that represents the nearest
blood-relatives of the vertebrates.
[ 192 ]
The discovery was confirmed by other zoologists, and
there can no longer be any doubt that of all the classes of
invertebrates that of the Tunicates is most closely related to the
vertebrates, and of the Tunicates the nearest are the
Ascidiæ. We cannot say that the vertebrates are descended
from the Ascidiæ—and still less the reverse—but
we can say that of all the invertebrates it is the Tunicates, and,
within this group, the Ascidiæ, that are the nearest
blood-relatives of the ancient stem-form of the vertebrates. We
must assume as the common ancestral group of both stems an extinct
family of the extensive vermalia-stem, the Prochordonia or
Prochordata (“primitive chorda-animals”).
In order to appreciate fully this remarkable fact, and
especially to secure the sound basis we seek for the genealogical
tree of the vertebrates, it is necessary to study thoroughly the
embryology of both these animals, and compare the individual
development of the Amphioxus step by step with that of the Ascidia.
We begin with the ontogeny of the Amphioxus.
From the concordant observations of Kowalevsky at Naples and
Hatschek at Messina, it follows, firstly, that the
ovum-segmentation and gastrulation of the Amphioxus are of the
simplest character. They take place in the same way as we find them
in many of the lower animals of different invertebrate stems, which
we have already described as original or primordial; the
development of the Ascidia is of the same type. Sexually mature
specimens of the Amphioxus, which are found in great quantities at
Messina from April or May onwards, begin as a rule to eject their
sexual products in the evening; if you catch them about the middle
of a warm night and put them in a glass vessel with seawater, they
immediately eject through the mouth their accumulated sexual
products, in consequence of the disturbance. The males give out
masses of sperm, and the females discharge ova in such quantity
that many of them stick to the fibrils about their mouths. Both
kinds of cells pass first into the mantle-cavity after the opening
of the gonads, proceed through the gill-clefts into the branchial
gut, and are discharged from this through the mouth.
The ova are simply round cells. They are only 1/250 of an inch
in diameter, and thus are only half the size of the mammal ova, and
have no distinctive features. The clear protoplasm of the mature
ovum is made so turbid by the numbers of dark granules of food-yelk
or deutoplasm scattered in it that it is difficult to follow the
process of fecundation and the behaviour of the two nuclei during
it (p. 51). The active elements of the
male sperm, the cone-shaped spermatozoa, are similar to those of
most other animals (cf. Fig. 20). Fecundation takes place when
these lively ciliated cells of the sperm approach the ovum, and
seek to penetrate into the yelk-matter or the cellular substance of
the ovum with their head-part—the thicker part of the cell
that encloses the nucleus. Only one spermatozoon can bore its way
into the yelk at one pole of the ovum-axis; its head or nucleus
coalesces with the female nucleus, which remains after the
extrusion of the directive bodies from the germinal vesicle. Thus
is formed the “stem-nucleus,” or the nucleus of the
“stem-cell” (cytula, Fig.
2). This now undergoes total segmentation, dividing into two,
four, eight, sixteen, thirty-two cells, and so on. In this way we
get the spherical, mulberry-shaped body, which we call the
morula.
The segmentation of the Amphioxus is not entirely regular, as
was supposed after the first observations of Kowalevsky (1866). It
is not completely equal, but a little unequal. As Hatschek
afterwards found (1879), the segmentation-cells only remain equal
up to the morula-stage, the spherical body of which consists of
thirty-two cells. Then, as always happens in unequal segmentation,
the more sluggish vegetal cells are outstripped in the cleavage. At
the lower or vegetal pole of the ovum a crown of eight large
entodermic cells remains for a long time unchanged, while the other
cells divide, owing to the formation of a series of horizontal
circles, into an increasing number of crowns of sixteen cells each.
Afterwards the segmentation-cells get more or less irregularly
displaced, while the segmentation-cavity enlarges in the centre of
the morula; in the end the former all lie on the surface of the
latter, so that the fœtus attains the familiar blastula shape
and forms a hollow ball, the wall of which consists of a single
stratum of cells (Fig. 38
A–C). This layer is the blastoderm, the simple
epithelium from the cells of which all the tissues of the body
proceed.
[ 193 ]
These important early embryonic processes take place so quickly
in the Amphioxus that four or five hours after fecundation, or
about midnight, the spherical blastula is completed. A pit-like
depression is then formed at the vegetal pole of it, and in
consequence of this the hollow sphere doubles on itself (Fig. 38
D). This pit becomes deeper and deeper (Fig. 38 E,
F); at last the invagination (or doubling) is complete, and the
inner or folded part of the blastula-wall lies on the inside of the
outer wall. We thus get a hollow hemisphere, the thin wall of which
is made up of two layers of cells (Fig. 38 E). From
hemispherical the body soon becomes almost spherical once more, and
then oval, the internal cavity enlarging considerably and its mouth
growing narrower (Fig. 213). The
form which the Amphioxus-embryo has thus reached is a real
“cup-larva” or gastrula, of the original simple
type that we have previously described as the
“bell-gastrula” or archigastrula (Figs.
29–35).
As in all the other animals that form an archigastrula, the
whole body is nothing but a simple gastric sac or stomach; its
internal cavity is the primitive gut (progaster or
archenteron, Fig. 38 g, 35 d), and its aperture
the primitive mouth (prostoma or blastoporus, o). The
wall is at once gut-wall and body-wall. It is composed of two
simple cell-layers, the familiar primary germinal layers. The inner
layer or the invaginated part of the blastoderm, which immediately
encloses the gut-cavity is the entoderm, the inner or vegetal
germ-layer, from which develop the wall of the alimentary canal and
all its appendages, the cœlom-pouches, etc. (Figs. 35, 36
i). The outer stratum of cells, or the non-invaginated part of
the blastoderm, is the ectoderm, the outer or animal germ-layer,
which provides the outer skin (epidermis) and the nervous system
(e). The cells of the entoderm are much larger, darker, and
more fatty than those of the ectoderm, which are clearer and less
rich in fatty particles. Hence before and during invagination there
is an increasing differentiation of the inner from the outer layer.
The animal cells of the outer layer soon develop vibratory hairs;
the vegetal cells of the inner layer do so much later. A
thread-like process grows out of each cell, and effects continuous
vibratory movements. By the vibrations of these slender hairs the
gastrula of the Amphioxus swims about in the sea, when it has
pierced the thin ovolemma, like the gastrula of many other animals
(Fig. 36). As in many other lower animals, the cells have only one
whip-like hair each, and so are called flagellate (whip)
cells (in contrast with the ciliated cells, which have a
number of short lashes or cilia).
In the further course of its rapid development the roundish
bell-gastrula becomes elongated, and begins to flatten on one side,
parallel to the long axis. The flattened side is the subsequent
dorsal side; the opposite or ventral side remains curved. The
latter grows more quickly than the former, with the result that the
primitive mouth is forced to the dorsal side (Fig. 39). In the
middle of the dorsal surface a shallow longitudinal groove or
furrow is formed (Fig. 79), and the edges of the body rise up on
each side of this groove in the shape of two parallel swellings.
This groove is, of course, the dorsal furrow, and the swellings are
the dorsal or medullary swellings; they form the first structure of
the central nervous system, the medullary tube. The medullary
swellings now rise higher; the groove between them becomes deeper
and deeper. The edges of the parallel swellings curve towards each
other, and at last unite, and the medullary tube is formed (Figs.
83 m, 84 m). Hence the formation of a medullary tube
out of the outer skin takes place in the naked dorsal surface of
the free-swimming larva of the Amphioxus in just the same way as we
have found in the embryo of man and the higher animals within the
fœtal membranes.
Simultaneously with the construction of the medullary tube we
have in the Amphioxus-embryo the formation of the chorda, the
cœlom-pouches, and the mesoderm proceeding from their wall.
These processes also take place with characteristic simplicity and
clearness, so that they are very instructive to compare with the
vermalia on the one hand and with the higher vertebrates on the
other. While the medullary groove is sinking in the middle line of
the flat dorsal side of the oval embryo, and its parallel edges
unite to form the ectodermic neural tube, the single chorda is
formed directly underneath them, and on each side of this a
parallel longitudinal fold, from the dorsal wall of the primitive
gut. These longitudinal folds of the entoderm proceed from the
primitive mouth, or from its lower
[ 194 ]
and hinder edge. Here we see at an early stage a
couple of large entodermic cells, which are distinguished from all
the others by their great size, round form, and fine-grained
protoplasm; they are the two promesoblasts, or polar cells of the
mesoderm (Fig. 83 p). They
indicate the original starting-point of the two
cœlom-pouches, which grow from this spot between the inner
and outer germinal layers, sever themselves from the primitive gut,
and provide the cellular material for the middle layer.
Immediately after their formation the two cœlom-pouches of
the Amphioxus are divided into several parts by longitudinal and
transverse folds. Each of the primary pouches is divided into an
upper dorsal and a lower ventral section by a couple of lateral
longitudinal folds (Fig. 82). But these are again divided by
several parallel transverse folds into a number of successive sacs,
the primitive segments or somites (formerly called by the
unsuitable name of “primitive vertebræ”). They
have a different future above and below. The upper or dorsal
segments, the episomites, lose their cavity later on, and
form with their cells the muscular plates of the trunk. The lower
or ventral segments, the hyposomites, corresponding to the
lateral plates of the craniote-embryo, fuse together in the upper
part owing to the disappearance of their lateral walls, and thus
form the later body-cavity (metacœl); in the lower part they
remain separate, and afterwards form the segmental gonads.
In the middle, between the two lateral cœlom-folds of the
primitive gut, a single central organ detaches from this at an
early stage in the middle line of its dorsal wall. This is the
dorsal chorda (Figs. 83, 84 ch). This axial rod, which is
the first foundation of the later vertebral column in all the
vertebrates, and is the only representative of it in the Amphioxus,
originates from the entoderm.
In consequence of these important folding-processes in the
primitive gut, the simple entodermic tube divides into four
different sections:— I, underneath, at the ventral side, the
permanent alimentary canal or permanent gut; II, above, at the
dorsal side, the axial rod or chorda; and III, the two
cœlom-sacs, which immediately sub-divide into two
structures:—IIIA, above, on the dorsal side,
the episomites, the double row of primitive or muscular
segments; and IIIB, below, on each side of the gut,
the hyposomites, the two lateral plates that give rise to
the sex-glands, and the cavities of which partly unite to form the
body-cavity. At the same time, the neural or medullary tube is
formed above the chorda, on the dorsal surface, by the closing of
the parallel medullary swellings. All these processes, which
outline the typical structure of the vertebrate, take place with
astonishing rapidity in the embryo of the Amphioxus; in the
afternoon of the first day, or twenty-four hours after
fertilisation, the young vertebrate, the typical embryo, is formed;
it then has, as a rule, six to eight somites.
The chief occurrence on the second day of development is the
construction of the two permanent openings of the gut—the
mouth and anus. In the earlier stages the alimentary tube is found
to be entirely closed, after the closing of the primitive mouth; it
only communicates behind by the neurenteric canal with the
medullary tube. The permanent mouth is a secondary formation, at
the opposite end. Here, at the end of the second day, we find a
pit-like depression in the outer skin, which penetrates inwards
into the closed gut. The anus is formed behind in the same way a
few hours later (in the vicinity of the additional gastrula-mouth).
In man and the higher vertebrates also the mouth and anus are
formed, as we have seen, as flat pits in the outer skin; they then
penetrate inwards, gradually becoming connected with the blind ends
of the closed gut-tube. During the second day the Amphioxus-embryo
undergoes few other changes. The number of primitive segments
increases, and generally amounts to fourteen, some forty-eight to
fifty hours after impregnation.
Almost simultaneously with the formation of the mouth the first
gill-cleft breaks through in the fore section of the
Amphioxus-embryo (generally forty hours after the commencement of
development). It now begins to nourish itself independently, as the
food material stored up in the ovum is completely used up. The
further development of the free larvæ takes place very
slowly, and extends over several months. The body becomes much
longer, and is compressed at the sides, the head-end being
broadened in a sort of triangle. Two rudimentary sense-organs are
developed in it. Inside we find the first blood-vessels, an upper
or dorsal vessel, corresponding to the aorta, between the gut and
the dorsal cord, and a lower or ventral
[ 195 ]
vessel, corresponding to the subintestinal vein, at
the lower border of the gut. Now, the gills or respiratory organs
also are formed at the fore-end of the alimentary canal. The whole
of the anterior or respiratory section of the gut is converted into
a gill-crate, which is pierced trellis-wise by numbers of
branchial-holes, as in the ascidia. This is done by the foremost
part of the gut-wall joining star-wise with the outer skin, and the
formation of clefts at the point of connection, piercing the wall
and leading into the gut from without. At first there are very few
of these branchial clefts; but there are soon a number of
them—first in one, then in two, rows. The foremost gill-cleft
is the oldest. In the end we have a sort of lattice work of fine
gill-clefts, supported on a number of stiff branchial rods; these
are connected in pairs by transverse rods.
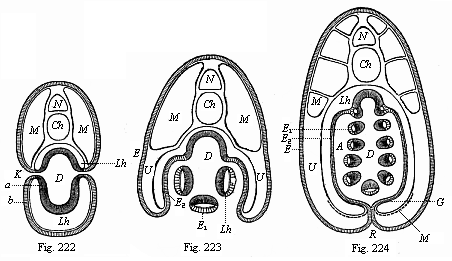
Figs. 222–224—Transverse
sections of young Amphioxus-larvæ (diagrammatic, from
Ralph.) (Cf. also Fig. 216.) In Fig. 222 there is free
communication from without with the gut-cavity (D) through
the gill-clefts (K). In Fig. 223 the lateral folds of the
body-wall, or the gill-covers, which grow downwards, are formed. In
Fig. 224 these lateral folds have united underneath and joined
their edges in the middle line of the ventral side (R seam).
The respiratory water now passes from the gut-cavity (D)
into the mantle-cavity (A). The letters have the same
meaning throughout: N medullary tube, Ch chorda,
M lateral muscles, Lh body-cavity, G part of the
body-cavity in which the sexual organs are subsequently formed.
D gut-cavity, clothed with the gut-gland layer (a). A
mantle-cavity, K gill-clefts, b=E epidermis,
E1 the same as visceral epithelium of the
mantle-cavity, E2 as parietal epithelium of the
mantle-cavity. |
At an early stage of embryonic development the structure of the
Amphioxus-larva is substantially the same as the ideal picture we
have previously formed of the “Primitive Vertebrate”
(Figs. 98–102). But the body afterwards undergoes various
modifications, especially in the fore-part. These modifications do
not concern us, as they depend on special adaptations, and do not
affect the hereditary vertebrate type. When the free-swimming
Amphioxus-larva is three months old, it abandons its pelagic habits
and changes into the young animal that lives in the sand. In spite
of its smallness (one-eighth of an inch), it has substantially the
same structure as the adult. As regards the remaining organs of the
Amphioxus, we need only mention that the gonads or sexual glands
are developed very late, immediately out of the inner cell-layer of
the
[ 196 ]
body-cavity. Although we can find afterwards no
continuation of the body-cavity (Fig. 216 U) in the lateral
walls of the mantle-cavity, in the gill-covers or mantle-folds
(Fig. 224 U), there is one present in the beginning (Fig.
224 Lh). The sexual cells are formed below, at the bottom of
this continuation (Fig. 224 S). For the rest, the subsequent
development into the adult Amphioxus of the larva we have followed
is so simple that we need not go further into it here.
We may now turn to the embryology of the Ascidia, an animal that
seems to stand so much lower and to be so much more simply
organised, remaining for the greater part of its life attached to
the bottom of the sea like a shapeless lump. It was a fortunate
accident that Kowalevsky first examined just those larger specimens
of the Ascidiæ that show most clearly the relationship of the
vertebrates to the invertebrates, and the larvæ of which
behave exactly like those of the Amphioxus in the first stages of
development. This resemblance is so close in the main features that
we have only to repeat what we have already said of the ontogenesis
of the Amphioxus.
The ovum of the larger Ascidia (Phallusia, Cynthia, etc.)
is a simple round cell of 1/250 to 1/125 of an inch in diameter. In
the thick fine-grained yelk we find a clear round germinal vesicle
of about 1/750 of an inch in diameter, and this encloses a small
embryonic spot or nucleolus. Inside the membrane that surrounds the
ovum, the stem-cell of the Ascidia, after fecundation, passes
through just the same metamorphoses as the stem-cell of the
Amphioxus. It undergoes total segmentation; it divides into two,
four, eight, sixteen, thirty-two cells, and so on. By continued
total cleavage the morula, or mulberry-shaped cluster of cells, is
formed. Fluid gathers inside it, and thus we get once more a
globular vesicle (the blastula); the wall of this is a single
stratum of cells, the blastoderm. A real gastrula (a simple
bell-gastrula) is formed from the blastula by invagination, in the
same way as in the amphioxus.
Up to this there is no definite ground in the embryology of the
Ascidiæ for bringing them into close relationship with the
Vertebrates; the same gastrula is formed in the same way in many
other animals of different stems. But we now find an embryonic
process that is peculiar to the Vertebrates, and that proves
irrefragably the affinity of the Ascidiæ to the Vertebrates.
From the epidermis of the gastrula a medullary tube is
formed on the dorsal side, and, between this and the primitive gut,
a chorda; these are the organs that are otherwise only found
in Vertebrates. The formation of these very important organs takes
place in the Ascidia-gastrula in precisely the same way as in that
of the Amphioxus. In the Ascidia (as in the other case) the oval
gastrula is first flattened on one side—the subsequent dorsal
side. A groove or furrow (the medullary groove) is sunk in the
middle line of the flat surface, and two parallel longitudinal
swellings arise on either side from the skin layer. These medullary
swellings join together over the furrow, and form a tube; in this
case, again, the neural or medullary tube is at first open in
front, and connected with the primitive gut behind by the
neurenteric canal. Further, in the Ascidia-larva also the two
permanent apertures of the alimentary canal only appear later, as
independent and new formations. The permanent mouth does not
develop from the primitive mouth of the gastrula; this primitive
mouth closes up, and the later anus is formed near it by
invagination from without, on the hinder end of the body, opposite
to the aperture of the medullary tube.
During these important processes, that take place in just the
same way in the Amphioxus, a tail-like projection grows out of the
posterior end of the larva-body, and the larva folds itself up
within the round ovolemma in such a way that the dorsal side is
curved and the tail is forced on to the ventral side. In this tail
is developed—starting from the primitive gut—a
cylindrical string of cells, the fore end of which pushes into the
body of the larva, between the alimentary canal and the neural
canal, and is no other than the chorda dorsalis. This important
organ had hitherto been found only in the Vertebrates, not a single
trace of it being discoverable in the Invertebrates. At first the
chorda only consists of a single row of large entodermic cells. It
is afterwards composed of several rows of cells. In the
Ascidia-larva, also, the chorda develops from the dorsal middle
part of the primitive gut, while the two cœlom-pouches detach
themselves from it on both sides. The simple body-cavity is formed
by the coalescence of the two.
When the Ascidia-larva has attained
[ 197 ]
this stage of development it begins to move about in
the ovolemma. This causes the membrane to burst. The larva emerges
from it, and swims about in the sea by means of its oar-like tail.
These free-swimming larvæ of the Ascidia have been known for
a long time. They were first observed by Darwin during his voyage
round the world in 1833. They resemble tadpoles in outward
appearance, and use their tails as oars, as the tadpoles do.
However, this lively and highly-developed condition does not last
long. At first there is a progressive development; the foremost
part of the medullary tube enlarges into a brain, and inside this
two single sense-organs are developed, a dorsal auditory vesicle
and a ventral eye. Then a heart is formed on the ventral side of
the animal, or the lower wall of the gut, in the same simple form
and at the same spot at which the heart is developed in man and all
the other vertebrates. In the lower muscular wall of the gut we
find a weal-like thickening, a solid, spindle-shaped string of
cells, which becomes hollow in the centre; it begins to contract in
different directions, now forward and now backward, as is the case
with the adult Ascidia. In this way the sanguineous fluid
accumulated in the hollow muscular tube is driven in alternate
directions into the blood-vessels, which develop at both ends of
the cardiac tube. One principal vessel runs along the dorsal side
of the gut, another along its ventral side. The former corresponds
to the aorta and the dorsal vessel in the worms. The other
corresponds to the subintestinal vein and the ventral vessel of the
worms.
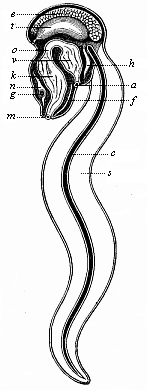 Fig.
225—An Appendicaria (Copelata), seen from the
left. m mouth, k branchial gut, o gullet,
v stomach, a anus, n brain (ganglion above the
gullet), g auditory vesicle, f ciliated groove under
the gills, h heart, t testicles, e ovary,
c chorda, s tail.
Fig.
225—An Appendicaria (Copelata), seen from the
left. m mouth, k branchial gut, o gullet,
v stomach, a anus, n brain (ganglion above the
gullet), g auditory vesicle, f ciliated groove under
the gills, h heart, t testicles, e ovary,
c chorda, s tail. |
With the formation of these organs the progressive development
of the Ascidia comes to an end, and degeneration sets in. The
free-swimming larva sinks to the floor of the sea, abandons its
locomotive habits, and attaches itself to stones, marine plants,
mussel-shells, corals, and other objects; this is done with the
part of the body that was foremost in movement. The attachment is
effected by a number of out-growths, usually three, which can be
seen even in the free-swimming larva. The tail is lost, as there is
no further use for it. It undergoes a fatty degeneration, and
disappears with the chorda dorsalis. The tailless body changes into
an unshapely tube, and, by the atrophy of some parts and the
modification of others, gradually assumes the appearance we have
already described.
Among the living Tunicates there is a very interesting group of
small animals that remain throughout life at the stage of
development of the tailed, free Ascidia-larva, and swim about
briskly in the sea by means of their broad oar-tail. These are the
remarkable Copelata (Appendicaria and Vexillaria,
Fig. 225). They are the only living Vertebrates that have
throughout life a chorda dorsalis and a neural string above it; the
latter must be regarded as the prolongation of the cerebral
ganglion and the equivalent of the medullary tube. Their branchial
gut also opens directly outwards by a pair of
[ 198 ]
branchial clefts. These instructive Copelata,
comparable to permanent Ascidia-larvæ, come next to the
extinct Prochordonia, those ancient worms which we must regard as
the common ancestors of the Tunicates and Vertebrates. The chorda
of the Appendicaria is a long, cylindrical string (Fig. 225
c), and serves as an attachment for the muscles that work the
flat oar-tail.
Among the various modifications which the Ascidia-larva
undergoes after its establishment at the sea-floor, the most
interesting (after the loss of the axial rod) is the atrophy of one
of its chief organs, the medullary tube. In the Amphioxus the
spinal marrow continues to develop, but in the Ascidia the tube
soon shrinks into a small and insignificant nervous ganglion that
lies above the mouth and the gill-crate, and is in accord with the
extremely slight mental power of the animal. This insignificant
relic of the medullary tube seems to be quite beyond comparison
with the nervous centre of the vertebrate, yet it started from the
same structure as the spinal cord of the Amphioxus. The
sense-organs that had been developed in the fore part of the neural
tube are also lost; no trace of which can be found in the adult
Ascidia. On the other hand, the alimentary canal becomes a most
extensive organ. It divides presently into two sections—a
wide fore or branchial gut that serves for respiration, and a
narrower hind or hepatic gut that accomplishes digestion. The
branchial or head-gut of the Ascidia is small at first, and opens
directly outwards only by a couple of lateral ducts or
gill-clefts—a permanent arrangement in the Copelata. The
gill-clefts are developed in the same way as in the Amphioxus. As
their number greatly increases we get a large gill-crate, pierced
like lattice work. In the middle line of its ventral side we find
the hypobranchial groove. The mantle or cloaca-cavity (the atrium)
that surrounds the gill-crate is also formed in the same way in the
Ascidia as in the Amphioxus. The ejection-opening of this
peribranchial cavity corresponds to the branchial pore of the
Amphioxus. In the adult Ascidia the branchial gut and the heart on
its ventral side are almost the only organs that recall the
original affinity with the vertebrates.
The further development of the Ascidia in detail has no
particular interest for us, and we will not go into it. The chief
result that we obtain from its embryology is the complete agreement
with that of the Amphioxus in the earliest and most important
embryonic stages. They do not begin to diverge until after the
medullary tube and alimentary canal, and the axial rod with the
muscles between the two, have been formed. The Amphioxus continues
to advance, and resembles the embryonic forms of the higher
vertebrates; the Ascidia degenerates more and more, and at last, in
its adult condition, has the appearance of a very imperfect
invertebrate.
If we now look back on all the remarkable features we have
encountered in the structure and the embryonic development of the
Amphioxus and the Ascidia, and compare them with the features of
man’s embryonic development which we have previously studied,
it will be clear that I have not exaggerated the importance of
these very interesting animals. It is evident that the Amphioxus
from the vertebrate side and the Ascidia from the invertebrate form
the bridge by which we can span the deep gulf that separates the
two great divisions of the animal kingdom. The radical agreement of
the lancelet and the sea-squirt in the first and most important
stages of development shows something more than their close
anatomic affinity and their proximity in classification; it shows
also their real blood-relationship and their common origin from one
and the same stem-form. In this way, it throws considerable light
on the oldest roots of man’s genealogical tree.
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XVI
Chapter XVIII
Figs. 1–209
Figs. 210–408