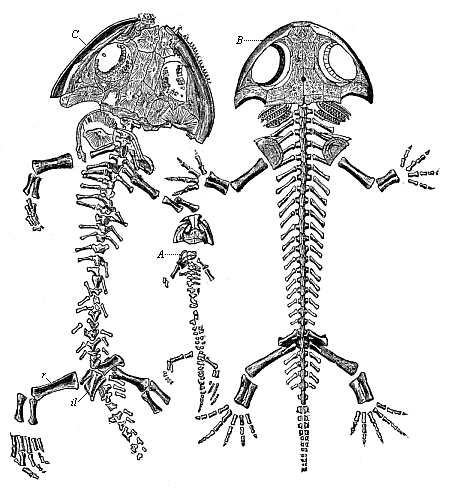
 Fig. 260—Fossil amphibian from the Permian, found in the Plauen terrain near Dresden (Branchiosaurus amblystomus). (From Credner.) A skeleton of a young larva. B larva, restored, with gills. C the adult form.)
Fig. 260—Fossil amphibian from the Permian, found in the Plauen terrain near Dresden (Branchiosaurus amblystomus). (From Credner.) A skeleton of a young larva. B larva, restored, with gills. C the adult form.)With the phylogenetic study of the four higher classes of Vertebrates, which must now engage our attention, we reach much firmer ground and more light in the construction of our genealogy than we have, perhaps, enjoyed up to the present. In the first place, we owe a number of very valuable data to the very interesting class of Vertebrates that come next to the Dipneusts and have been developed from them—the Amphibia. To this group belong the salamander, the frog, and the toad. In earlier days all the reptiles were, on the example of Linne, classed with the Amphibia (lizards, serpents, crocodiles, and tortoises). But the reptiles are much more advanced than the Amphibia, and are nearer to the birds in the chief points of their structure. The true Amphibia are nearer to the Dipneusta and the fishes; they are also much older than the reptiles. There were plenty of highly-developed (and sometimes large) Amphibia during the Carboniferous period; but the earliest reptiles are only found in the Permian period. It is probable that the Amphibia were evolved even earlier—during the Devonian period—from the Dipneusta. The extinct Amphibia of which we have fossil remains from that remote period (very numerous especially in the Triassic strata) were distinguished for a graceful scaly coat or a powerful bony armour on the skin (like the crocodile), whereas the living amphibia have usually a smooth and slippery skin.
The earliest of these armoured Amphibia (Phractamphibia) form the order of Stegocephala (“roof-headed”) (Fig. 260). It is among these, and not among the actual Amphibia, that we must look for the forms that are directly related to the genealogy of our race, and are the ancestors of the three higher classes of Vertebrates. But even the existing Amphibia have such important relations to us in their anatomic structure, and especially their embryonic development, that we may say: Between the Dipneusts and the Amniotes there was a series of extinct intermediate forms which we should certainly class with the Amphibia if we had them before us. In their whole organisation even the actual Amphibia seem to be an instructive transitional group. In the important respects of respiration and circulation they approach very closely to the Dipneusta, though in other respects they are far superior to them.
This is particularly true of the development of their limbs or extremities. In them we find these for the first time as five-toed feet. The thorough investigations of Gegenbaur have shown that the fish’s fins, of which very erroneous opinions were formerly held, are many-toed feet. The various cartilaginous or bony radii that are found in large numbers in each fin correspond to the fingers or toes of the higher Vertebrates. The several joints of each fin-radius correspond to the various parts of the toe. Even in the Dipneusta the fin is of the same construction as in the fishes; it was afterwards gradually evolved into the five-toed form, which we first encounter in the Amphibia. This reduction of the number of the toes to six, and then to five, probably took place in the second half of the Devonian period—at the latest, in the subsequent Carboniferous period—in those Dipneusta which we regard as the ancestors of the Amphibia. We have several fossil remains of five-toed Amphibia from this period. There are numbers of fossil impressions of them in the Triassic of Thuringia (Chirotherium).
|
Fig. 260—Fossil amphibian from the Permian, found in the Plauen terrain near Dresden (Branchiosaurus amblystomus). (From Credner.) A skeleton of a young larva. B larva, restored, with gills. C the adult form.) |
The fact that the toes number five is of great importance, because they have clearly been transmitted from the Amphibia to all the higher Vertebrates. Man entirely resembles his amphibian ancestors in this respect, and indeed in the whole structure of the bony skeleton of his five-toed extremities. A careful comparison of the skeleton of the frog with our own is enough to show this. It is well known that this hereditary number of the toes has assumed a very great practical importance from remote times; on it our whole system of enumeration (the decimal system applied to measurement of time, mass, weight, etc.) is based. There is absolutely no reason why there should be five toes in the fore and hind feet in the lowest Amphibia, the reptiles, and the higher Vertebrates, unless we
ascribe it to inheritance from a common stem-form. Heredity alone can explain it. It is true that we find less than five toes in many of the Amphibia and of the higher Vertebrates. But in all these cases we can prove that some of the toes atrophied, and were in time lost altogether.
The causes of this evolution of the five-toed foot from the many-toed fin in the amphibian ancestor must be sought in adaptation to the entire change of function that the limbs experienced in passing from an exclusively aquatic to a partly terrestrial life. The many-toed fin had been used almost solely for motion in the water; it had now also to support the body in creeping on the solid ground. This led to a modification both of the skeleton and the muscles of the limbs. The number of the fin-radii was gradually reduced, and sank finally to five. But these five remaining radii became much stronger. The soft cartilaginous radii became bony rods. The rest of the skeleton was similarly strengthened. Thus from the one-armed lever of the many-toed fish-fin arose the improved many-armed lever system of the five-toed amphibian limbs. The movements of the body gained in variety as well as in strength. The various parts of the skeletal system and correlated muscular system began to differentiate more and more. In view of the close correlation of the muscular and nervous systems, this also made great advance in structure and function. Hence we find, as a matter of fact, that the brain is much more developed in the higher Amphibia than in the fishes, the Dipneusta, and the lower Amphibia.
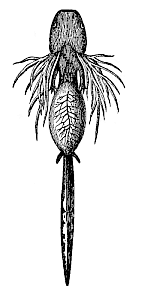 Fig. 261—Larva of the Spotted Salamander (Salamandra maculata), seen from the ventral side. In the centre a yelk-sac still hangs from the gut. The external gills are gracefully ramified. The two pairs of legs are still very small.
Fig. 261—Larva of the Spotted Salamander (Salamandra maculata), seen from the ventral side. In the centre a yelk-sac still hangs from the gut. The external gills are gracefully ramified. The two pairs of legs are still very small. |
The first advance in organisation that was occasioned by the adoption of life on land was naturally the construction of an organ for breathing air—a lung. This was formed directly from the floating-bladder inherited from the fishes. At first its function was insignificant beside that of the gills, the older organ for water-respiration. Hence we find in the lowest Amphibia, the gilled Amphibia, that, like the Dipneusta, they pass the greater part of their life in the water, and breathe water through gills. They only come to the surface at brief intervals, or creep on to the land, and then breathe air by their lungs. But some of the tailed Amphibia—the salamanders—remain entirely in the water when they are young, and afterwards spend most of their time on land. In the adult state they only breathe air through lungs. The same applies to the most advanced of the Amphibia, the Batrachia (frogs and toads); some of them have entirely lost the gill-bearing larva form.1 This is also the case with certain small, serpentine Amphibia, the Cæcilia (which live in the ground like earth-worms).
The great interest of the natural history of the Amphibia consists especially in their intermediate position between the lower and higher Vertebrates. The lower Amphibia approach very closely to the Dipneusta in their whole organisation, live mainly in the water, and breathe by gills; but the higher Amphibia are just as close to the Amniotes, live mainly on land, and breathe by lungs. But in their younger state the latter resemble the former, and only reach the higher stage by a complete metamorphosis. The embryonic development of most of the
1. The tree-frog of Martinique (Hylades martinicensis) loses the gills on the seventh, and the tail and yelk-sac on the eighth, day of fœtal life. On the ninth or tenth day after fecundation the frog emerges from the egg.
higher Amphibia still faithfully reproduces the stem-history of the whole class, and the various stages of the advance that was made by the lower Vertebrates in passing from aquatic to terrestrial life during the Devonian or the Carboniferous period are repeated in the spring by every frog that develops from an egg in our ponds.
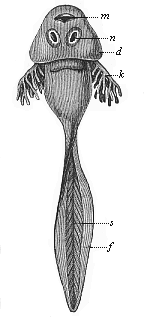 Fig. 262—Larva of the common grass-frog (Rana temporaria), or “tadpole.” m mouth, n a pair of suckers for fastening on to stones, d skin-fold from which the gill-cover develops; behind it the gill-clefts, from which the branching gills (k) protrude, s tail-muscles, f cutaneous fin-fringe of the tail.
Fig. 262—Larva of the common grass-frog (Rana temporaria), or “tadpole.” m mouth, n a pair of suckers for fastening on to stones, d skin-fold from which the gill-cover develops; behind it the gill-clefts, from which the branching gills (k) protrude, s tail-muscles, f cutaneous fin-fringe of the tail. |
The common frog leaves the egg in the shape of a larva, like the tailed salamander (Fig. 261), and this is altogether different from the mature frog (Fig. 262). The short trunk ends in a long tail, with the form and structure of a fish’s tail (s). There are no limbs at first. The respiration is exclusively branchial, first through external (k) and then internal gills. In harmony with this the heart has the same structure as in the fish, and consists of two sections—an atrium that receives the venous blood from the body, and a ventricle that forces it through the arteries into the gills.
We find the larvæ of the frog (or tadpoles, Gyrini) in great numbers in our ponds every spring in this fish-form, using their muscular tails in swimming, just like the fishes and young Ascidia. When they have reached a certain size, the remarkable metamorphosis from the fish-form to the frog begins. A blind sac grows out of the gullet, and expands into a couple of spacious sacs: these are the lungs. The simple chamber of the heart is divided into two sections by the development of a partition, and there are at the same time considerable changes in the structure of the chief arteries. Previously all the blood went from the auricle through the aortic arches into the gills, but now only part of it goes to the gills, the other part passing to the lungs through the new-formed pulmonary artery. From this point arterial blood returns to the left auricle of the heart, while the venous blood gathers in the right auricle. As both auricles open into a single ventricle, this contains mixed blood. The dipneust form has now succeeded to the fish-form. In the further course of the metamorphosis the gills and the branchial vessels entirely disappear, and the respiration becomes exclusively pulmonary. Later, the long swimming tail is lost, and the frog now hops to the land with the legs that have grown meantime.
This remarkable metamorphosis of the Amphibia is very instructive in connection with our human genealogy, and is particularly interesting from the fact that the various groups of actual Amphibia have remained at different stages of their stem-history, in harmony with the biogenetic law. We have first of all a very low order of Amphibia—the Sozobranchia (“gilled-amphibia”), which retain their gills throughout life, like the fishes. In a second order of the salamanders the gills are lost in the metamorphosis, and when fully grown they have only pulmonary respiration. Some of the tailed Amphibia still retain the gill-clefts in the side of the neck, though they have lost the gills themselves (Menopoma). If we force the larvæ of our salamanders (Fig. 261) and tritons to remain in the water, and prevent them from reaching the land, we can in favourable circumstances make them retain their gills. In this fish-like condition they reach sexual maturity, and remain throughout life at the lower stage of the gilled Amphibia.
We have the reverse of this experiment in a Mexican gilled salamander, the fish-like axolotl (Siredon pisciformis). It was formerly regarded as a permanent gilled amphibian persisting throughout life at the fish-stage. But some of the hundreds of these animals that are kept in the Botanical Garden at Paris got on to the land for some reason or other, lost their gills, and changed into a form closely resembling the salamander (Amblystoma). Other species of the genus became sexually mature for the first time in this condition. This has been regarded as an astounding phenomenon, although every common frog and salamander repeats the metamorphosis in the spring. The whole change from the aquatic and gill-breathing animal to the terrestrial lung-breathing form may be followed step by step in this case. But what we see here in the development of the individual has happened to the whole class in the course of its stem-history.
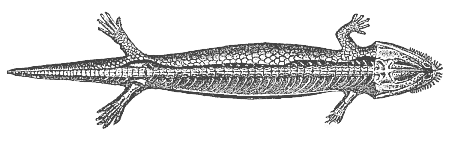 Fig. 263—Fossil mailed amphibian, from the Bohemian Carboniferous (Seeleya). (From Fritsch.) The scaly coat is retained on the left.
Fig. 263—Fossil mailed amphibian, from the Bohemian Carboniferous (Seeleya). (From Fritsch.) The scaly coat is retained on the left. |
The metamorphosis goes farther in a third order of Amphibia, the Batrachia or Anura, than in the salamander. To this belong the various kinds of toads, ringed snakes, water-frogs, tree-frogs, etc. These lose, not only the gills, but also (sooner or later) the tail, during metamorphosis.
The ontogenetic loss of the gills and the tail in the frog and toad can only be explained on the assumption that they are descended from long-tailed Amphibia of the salamander type. This is also clear from the comparative anatomy of the two groups. This remarkable metamorphosis is, however, also interesting because it throws a certain light on the phylogeny of the tail-less apes and man. Their ancestors also had long tails and gills like the gilled Amphibia, as the tail and the gill-arches of the human embryo clearly show.
For comparative anatomical and ontogenetic reasons, we must not seek these amphibian ancestors of ours—as one would be inclined to do, perhaps—among the tail-less Batrachia, but among the tailed lower Amphibia.
The vertebrate form that comes next to the Amphibia in the series of our ancestors is a lizard-like animal, the earlier existence of which can be confidently deduced from the facts of comparative anatomy and ontogeny. The living Hatteria of New Zealand (Fig. 264) and the extinct Rhyncocephala of the Permian period (Fig. 265) are closely related to this important stem-form; we may call them the Protamniotes, or Primitive Amniotes. All the Vertebrates above the Amphibia—or the three classes of reptiles, birds, and mammals—differ so much in their whole organisation from all the lower Vertebrates we have yet considered, and have so great a resemblance to each other, that we put them all together in a single group with the title of Amniotes. In these three classes alone we find the remarkable embryonic membrane, already mentioned, which we called the amnion; a cenogenetic adaptation that we may regard as a result of the sinking of the growing embryo into the yelk-sac.
All the Amniotes known to us—all reptiles, birds, and mammals (including man)—agree in so many important points of internal structure and development that their descent from a common ancestor can be affirmed with tolerable certainty. If the evidence of comparative
anatomy and ontogeny is ever entirely beyond suspicion, it is certainly the case here. All the peculiarities that accompany and follow the formation of the amnion, and that we have learned in our consideration of human embryology; all the peculiarities in the development of the organs which we will presently follow in detail; finally, all the principal special features of the internal structure of the full-grown Amniotes—prove so clearly the common origin of all the Amniotes from single extinct stem-form that it is difficult to entertain the idea of their evolution from several independent stems. This unknown common stem-form is our primitive Amniote (Protamnion). In outward appearance it was probably something between the salamander and the lizard.
It is very probable that some part of the Permian period was the age of the origin of the Protamniotes. This follows from the fact that the Amphibia are not fully developed until the Carboniferous period, and that the first fossil reptiles (Palæhatteria, Homœosaurus, Proterosaurus) are found towards the close of the Permian period. Among the important changes of the vertebrate organisation that marked the rise of the first Amniotes from salamandrine Amphibia during this period the following three are especially noteworthy: the entire disappearance of the water-breathing gills and the conversion of the gill-arches into other organs, the formation of the allantois or primitive urinary sac, and the development of the amnion.
One of the most salient characteristics of the Amniotes is the complete loss of the gills. All Amniotes, even if living in water (such as sea-serpents and whales), breathe air through lungs, never water through gills. All the Amphibia (with very rare exceptions) retain their gills for some time when young, and have for a time (if not permanently) branchial respiration; but after these there is no question of branchial respiration. The Protamniote itself must have entirely abandoned water-breathing. Nevertheless, the gill-arches are preserved by heredity, and develop into totally different (in part rudimentary) organs—various parts of the bone of the tongue, the frame of the jaws, the organ of hearing, etc. But we do not find in the embryos of the Amniotes any trace of gill-leaves, or of real respiratory organs on the gill-arches.
With this complete abandonment of the gills is probably connected the formation of another organ, to which we have already referred in embryology—namely, the allantois or primitive urinary sac (cf. p. 166). It is very probable that the urinary bladder of the Dipneusts is the first structure of the allantois. We find in these a urinary bladder that proceeds from the lower wall of the hind end of the gut, and serves as receptacle for the renal secretions. This organ has been transmitted to the Amphibia, as we can see in the frog.
The formation of the amnion and the allantois and the complete disappearance of the gills are the chief characteristics that distinguish the Amniotes from the lower Vertebrates we have hitherto considered. To these we may add several subordinate features that are transmitted to all the Amniotes, and are found in these only. One striking embryonic character of the Amniotes is the great curve of the head and neck in the embryo. We also find an advance in the structure of several of the internal organs of the Amniotes which raises them above the highest of the anamnia. In particular, a partition is formed in the simple ventricle of the heart, dividing into right and left chambers. In connection with the complete metamorphosis of the gill-arches we find a further development of the auscultory organs. Also, there is a great advance in the structure of the brain, skeleton, muscular system, and other parts. Finally, one of the most important changes is the reconstruction of the kidneys. In all the earlier Vertebrates we have found the primitive kidneys as excretory organs, and these appear at an early stage in the embryos of all the higher Vertebrates up to man. But in the Amniotes these primitive kidneys cease to act at an early stage of embryonic life, and their function is taken up by the permanent or secondary kidneys, which develop from the terminal section of the prorenal ducts.
Taking all these peculiarities of the Amniotes together, it is impossible to doubt that all the animals of this group—all reptiles, birds, and mammals—have a common origin, and form a single blood-related stem. Our own race belongs to this stem. Man is, in every feature of his organisation and embryonic development, a true Amniote, and has descended from the Protamniote with all the other
Amniotes. Though they appeared at the end (possibly even in the middle) of the Paleozoic age, the Amniotes only reached their full development during the Mesozoic age. The birds and mammals made their first appearance during this period. Even the reptiles show their greatest growth at this time, so that it is called “the reptile age.” The extinct Protamniote, the ancestor of the whole group, belongs in its whole organisation to the reptile class.
 Fig. 264—The lizard (Hatteria punctata = Sphenodon punctatus) of New Zealand. The sole surviving proreptile. (From Brehm.)
Fig. 264—The lizard (Hatteria punctata = Sphenodon punctatus) of New Zealand. The sole surviving proreptile. (From Brehm.) |
The genealogical tree of the amniote group is clearly indicated in its chief lines by their paleontology, comparative anatomy, and ontogeny. The group succeeding the Protamniote divided into two branches. The branch that will claim our whole interest is the class of the Mammals. The other branch, which developed in a totally different direction, and only comes in contact with the Mammals at its root, is the combined group of the reptiles and birds; these two classes may, with Huxley, be conveniently grouped together as the Sauropsida. Their common stem-form is an extinct lizard-like reptile of the order of the Rhyncocephalia. From this have been developed in various directions the serpents, crocodiles, tortoises, etc.—in a word, all the members of the reptile class. But the remarkable class of the birds has also been evolved directly from a branch of the reptile group, as is now established beyond question. The embryos of the reptiles and birds are identical until a very late stage, and have an astonishing resemblance even later. Their whole structure agrees so much that no anatomist now questions the descent of the birds from the reptiles. On the other
hand, the mammal line has descended from the group of the Sauromammalia, a different branch of the Proreptilia. It is connected at its deepest roots with the reptile line, but it then diverges completely from it and follows a distinctive development. Man is the highest outcome of this class, the “crown of creation.” The hypothesis that the three higher Vertebrate classes represent a single Amniote-stem, and that the common root of this stem is to be found in the amphibian class, is now generally admitted.
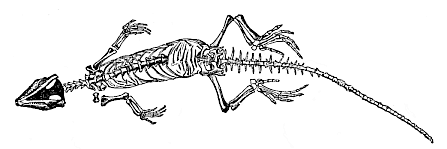 Fig. 265—Homœosaurus pulchellus, a Jurassic proreptile from Kehlheim. (From Zittel.)
Fig. 265—Homœosaurus pulchellus, a Jurassic proreptile from Kehlheim. (From Zittel.) |
The instructive group of the Permian Tocosauria, the common root from which the divergent stems of the Sauropsids and mammals have issued, merits our particular attention as the stem-group of all the Amniotes. Fortunately a living representative of this extinct ancestral group has been preserved to our day; this is the remarkable lizard of New Zealand, Hatteria punctata (Fig. 264). Externally it differs little from the ordinary lizard; but in many important points of internal structure, especially in the primitive construction of the vertebral column, the skull, and the limbs, it occupies a much lower position, and approaches its amphibian ancestors, the Stegocephala. Hence Hatteria is the phylogenetically oldest of all living reptiles, an isolated survivor from the Permian period, closely resembling the common ancestor of the Amniotes. It must differ so little from this extinct form, our hypothetical Protamniote, that we put it next to the Proreptilia. The remarkable Permian Palæhatteria, that Credner discovered in the Plauen terrain at Dresden in 1888, belongs to the same group (Fig. 266). The Jurassic genus Homœosaurus (Fig. 265), of which well-preserved skeletons are found in the Solenhofen schists, is perhaps still more closely related to them.
Unfortunately, the numerous fossil remains of Permian and Triassic Tocosauria that we have found in the last two decades are, for the most part, very imperfectly preserved. Very often we can make only precarious inferences from these skeletal fragments as to the anatomic characters of the soft parts that went with the bony skeleton of the extinct Tocosauria. Hence it has not yet been possible to arrange these important fossils with any confidence in the ancestral series that descend from the Protamniotes to the Sauropsids on the one side and the Mammals on the other. Opinions are particularly divided as to the place in classification and the phylogenetic significance of the remarkable Theromorpha. Cope gives this name to a very interesting and extensive group of extinct terrestrial reptiles, of which we have only fossil remains from the Permian and Triassic strata. Forty years ago some of these Therosauria (fresh-water animals) were described by Owen as Anomodontia. But during the last twenty years the distinguished American paleontologists, Cope and Osborn, have greatly increased our knowledge of them, and have claimed that the stem-forms of the Mammals must be sought in this order. As a matter of fact, the Theromorpha are nearer to the Mammals in the chief points of structure than any other reptiles. This is especially true of the Thereodontia, to which the Pureosauria and Pelycosauria belong (Fig. 267). The whole structure of their pelvis and hind-feet has attained the same form as in the Monotremes, the lowest Mammals. The formation of the
scapula and the quadrate bone shows an approach to the Mammals such as we find in no other group of reptiles. The teeth also are already divided into incisors, canines, and molars. Nevertheless, it is very doubtful whether the Theromorpha really are in the ancestral line of the Sauromammals, or lead direct from the Tocosauria to the earliest Mammals. Other experts on this group believe that it is an independent legion of the reptiles, connected, perhaps, at its lowest root, with the Sauromammals, but developed quite independently of the Mammals—though parallel to them in many ways.
One of the most important of the zoological facts that we rely on in our investigation of the genealogy of the human race is the position of man in the Mammal class. However different the views of zoologists may have been as to this position in detail, and as to his relations to the apes, no scientist has ever doubted that man is a true mammal in his whole organisation and development. Linné drew attention to this fact in the first edition of his famous Systema Naturæ (1735). As will be seen in any museum of anatomy or any manual of comparative anatomy; the human frame has all the characteristics that are common to the Mammals and distinguish them conspicuously from all other animals.
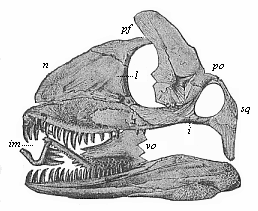 Fig. 266—Skull of a Permian lizard (Palæhatteria longicaudata). (From Credner.) n nasal bone, pf frontal bone, l lachrymal bone, po postorbital bone, sq covering bone, i cheek-bone, vo vomer, im inter-maxillary.
Fig. 266—Skull of a Permian lizard (Palæhatteria longicaudata). (From Credner.) n nasal bone, pf frontal bone, l lachrymal bone, po postorbital bone, sq covering bone, i cheek-bone, vo vomer, im inter-maxillary. |
If we examine this undoubted fact from the point of view of phylogeny, in the light of the theory of descent, it follows at once that man is of a common stem with all the other Mammals, and comes from the same root as they. But the various features in which the Mammals agree and by which they are distinguished are of such a character as to make a polyphyletic hypothesis quite inadmissible. It is impossible to entertain the idea that all the living and extinct Mammals come from a number of separate roots. If we accept the general theory of evolution, we are bound to admit the monophyletic hypothesis of the descent of all the Mammals (including man) from a single mammalian stem-form. We may call this long-extinct root-form and its earliest descendants (a few genera of one family) “primitive mammals” or “stem-mammals” (Promammalia). As we have already seen, this root-form developed from the primitive Proreptile stem in a totally different direction from the birds, and soon separated from the main stem of the reptiles. The differences between the Mammals and the reptiles and birds are so important and characteristic that we can assume with complete confidence this division of the vertebrate stem at the commencement of the development of the Amniotes. The reptiles and birds, which we group together as the Sauropsids, generally agree in the characteristic structure of the skull and brain, and this is notably different from that of the Mammals. In most of the reptiles and birds the skull is connected with the first cervical vertebra (the atlas) by a single, and in the Mammals (and Amphibia) by a double, condyle at the back of the head. In the former the lower jaw is composed of several pieces, and connected with the skull so that it can move by a special maxillary bone (the quadratum); in the Mammals the lower jaw consists of one pair of bony pieces, which articulate directly with the temporal bone. Further, in the Sauropsids the skin is clothed with scales or feathers; in the Mammals with hair. The red blood-cells of the former have a nucleus; those of the latter have not. In fine, two quite characteristic features of the Mammals, which distinguish them not only from the birds and reptiles, but from all other animals, are the possession of a
complete diaphragm and of mammary glands that produce the milk for the nutrition of the young. It is only in the Mammals that the diaphragm forms a transverse partition of the body-cavity, completely separating the pectoral from the abdominal cavity. It is only in the mammals that the mother suckles its young, and this rightly gives the name to the whole class (mamma = breast).
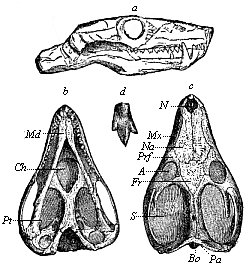 Fig. 267—Skull of a Triassic theromorphum (Galesaurus planiceps), from the Karoo formation in South Africa. (From Owen.) a from the right, b from below, c from above, d tricuspid tooth. N nostrils, Na nasal bone, Mx upper jaw, Prf prefrontal, Fr frontal bone, A eye-pits, S temple-pits. Pa Parietal eye, Bo joint at back of head, Pt pterygoid-bone, Md lower jaw.
Fig. 267—Skull of a Triassic theromorphum (Galesaurus planiceps), from the Karoo formation in South Africa. (From Owen.) a from the right, b from below, c from above, d tricuspid tooth. N nostrils, Na nasal bone, Mx upper jaw, Prf prefrontal, Fr frontal bone, A eye-pits, S temple-pits. Pa Parietal eye, Bo joint at back of head, Pt pterygoid-bone, Md lower jaw. |
From these pregnant facts of comparative anatomy and ontogeny it follows absolutely that the whole of the Mammals belong to a single natural stem, which branched off at an early date from the reptile-root. It follows further with the same absolute certainty that the human race is also a branch of this stem. Man shares all the characteristics I have described with all the Mammals, and differs in them from all other animals. Finally, from these facts we deduce with the same confidence those advances in the vertebrate organisation by which one branch of the Sauromammals was converted into the stem-form of the Mammals. Of these advances the chief were: (1) The characteristic modification of the skull and the brain; (2) the development of a hairy coat; (3) the complete formation of the diaphragm; and (4) the construction of the mammary glands and adaptation to suckling. Other important changes of structure proceeded step by step with these.
The epoch at which these important advances were made, and the foundation of the Mammal class was laid, may be put with great probability in the first section of the Mesozoic or secondary age—the Triassic period. The oldest fossil remains of mammals that we know were found in strata that belong to the earliest Triassic period—the upper Kueper. One of the earliest forms is the genus Dromatherium, from the North American Triassic (Fig. 268). Their teeth still strikingly recall those of the Pelycosauria. Hence we may assume that this small and probably insectivorous mammal belonged to the stem-group of the Promammals. We do not find any positive trace of the third and most advanced division of the Mammals—the Placentals. These (including man) are much younger, and we do not find indisputable fossil remains of them until the Cenozoic age, or the Tertiary period. This paleontological fact is very important, because it fully harmonises with the evolutionary succession of the Mammal orders that is deduced from their comparative anatomy and ontogeny.
The latter science teaches us that the whole Mammal class divides into three main groups or sub-classes, which correspond to three successive phylogenetic stages. These three stages, which also represent three important stages in our human genealogy, were first distinguished in 1816 by the eminent French zoologist, Blainville, and received the names of Ornithodelphia, Didelphia, and Monodelphia, according to the construction of the female organs (delphys = uterus or womb). Huxley afterwards gave them the names of Prototheria, Metatheria, and Epitheria. But the three sub-classes differ so widely from each other, not only in the construction of the sexual organs, but in many other respects also, that we may confidently draw up the following
important phylogenetic thesis: The Monodelphia or Placentals descend from the Didelphia or Marsupials; and the latter, in turn, are descended from the Monotremes or Ornithodelphia.
Thus we must regard as the twenty-first stage in our genealogical tree the earliest and lowest chief group of the Mammals—the sub-class of the Monotremes (“cloaca-animals,” Ornithodelphia, or Prototheria, Figs. 269 and 270). They take their name from the cloaca which they share with all the lower Vertebrates. This cloaca is the common outlet for the passage of the excrements, the urine, and the sexual products. The urinary ducts and sexual canals open into the hindmost part of the gut, while in all the other Mammals they are separated from the rectum and anus. The latter have a special uro-genital outlet (porus urogenitalis). The bladder also opens into the cloaca in the Monotremes, and, indeed, apart from the two urinary ducts; in all the other Mammals the latter open directly into the bladder. It was proved by Haacke and Caldwell in 1884 that the Monotremes lay large eggs like the reptiles, while all the other Mammals are viviparous. In 1894 Richard Semon further proved that these large eggs, rich in food-yelk, have a partial segmentation and discoid gastrulation, as I had hypothetically assumed in 1879; here again they resemble their reptilian ancestors. The construction of the mammary gland is also peculiar in the Monotremes. In them the glands have no teats for the young animal to suck, but there is a special part of the breast pierced with holes like a sieve, from which the milk issues, and the young Monotreme must lick it off. Further, the brain of the Monotremes is very little advanced. It is feebler than that of any of the other Mammals. The fore-brain or cerebrum, in particular, is so small that it does not cover the cerebellum. In the skeleton (Fig. 270) the formation of the scapula among other parts is curious; it is quite different from that of the other Mammals, and rather agrees with that of the reptiles and Amphibia. Like these, the Monotremes have a strongly developed caracoideum. From these and other less prominent characteristics it follows absolutely that the Monotremes occupy the lowest place among the Mammals, and represent a transitional group between the Tocosauria and the rest of the Mammals. All these remarkable reptilian characters must have been possessed by the stem-form of the whole mammal class, the Promammal of the Triassic period, and have been inherited from the Proreptiles.
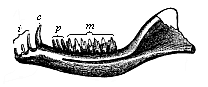 Fig. 268—Lower jaw of a Primitive Mammal or Promammal (Dromatherium silvestre) from the North American Triassic. i incisors, c canine, p premolars, m molars. (From Döderlein.)
Fig. 268—Lower jaw of a Primitive Mammal or Promammal (Dromatherium silvestre) from the North American Triassic. i incisors, c canine, p premolars, m molars. (From Döderlein.) |
During the Triassic and Jurassic periods the sub-class of the Monotremes was represented by a number of different stem-mammals. Numerous fossil remains of them have lately been discovered in the Mesozoic strata of Europe, Africa, and America. To-day there are only two surviving specimens of the group, which we place together in the family of the duck-bills, Ornithostoma. They are confined to Australia and the neighbouring island of Van Diemen’s Land (or Tasmania); they become scarcer every year, and will soon, like their blood-relatives, be counted among the extinct animals. One form lives in the rivers, and builds subterraneous dwellings on the banks; this is the Ornithorhyncus paradoxus, with webbed feet, a thick soft fur, and broad flat jaws, which look very much like the bill of a duck (Figs. 269, 270). The other form, the land duck-bill, or spiny ant-eater (Echidna hystrix), is very much like the anteaters in its habits and the peculiar construction of its thin snout and very long tongue; it is covered with needles, and can roll itself up like a hedgehog. A cognate form (Parechidna Bruyni) has lately been found in New Guinea.
These modern Ornithostoma are the scattered survivors of the vast Mesozoic group of Monotremes; hence they have the same interest in connection with the stem history of the Mammals as the living stem-reptiles (Hatteria) for that of the reptiles, and the isolated Acrania (Amphioxus) for the phylogeny of the Vertebrate stem.
The Australian duck-bills are distinguished externally by a toothless bird-like
beak or snout. This absence of real bony teeth is a late result of adaptation, as in the toothless Placentals (Edentata, armadillos and ant-eaters). The extinct Monotremes, to which the Promammalia belonged, must have had developed teeth, inherited from the reptiles. Lately small rudiments of real molars have been discovered in the young of the Ornithorhyncus, which has horny plates in the jaws instead of real teeth.
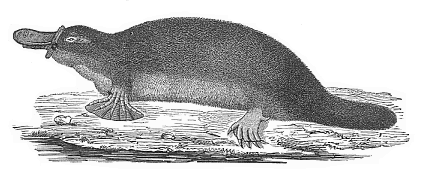 Fig. 269—The Ornithorhyncus or Duck-mole. (Ornithorhyncus paradoxus).
Fig. 269—The Ornithorhyncus or Duck-mole. (Ornithorhyncus paradoxus).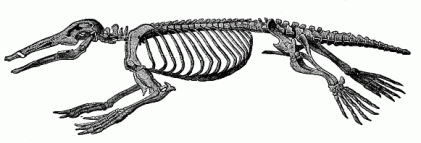 Fig. 270—Skeleton of the Ornithorhyncus.
Fig. 270—Skeleton of the Ornithorhyncus. |
The living Ornithostoma and the stem-forms of the Marsupials (or Didelphia) must be regarded as two widely diverging lines from the Promammals. This second sub-class of the Mammals is very interesting as a perfect intermediate stage between the other two. While the Marsupials retain a great part of the characteristics of the Monotremes, they have also acquired some of the chief features of the Placentals. Some features
are also peculiar to the Marsupials, such as the construction of the male and female sexual organs and the form of the lower jaw. The Marsupials are distinguished by a peculiar hook-like bony process that bends from the corner of the lower jaw and points inwards. As most of the Placentals have not this process, we can, with some probability, recognise the Marsupial from this feature alone. Most of the mammal remains that we have from the Jurassic and Cretaceous deposits are merely lower jaws, and most of the jaws found in the Jurassic deposits at Stonesfield and Purbeck have the peculiar hook-like process that characterises the lower jaw of the Marsupial. On the strength of this paleontological fact, we may suppose that they belonged to Marsupials. Placentals do not seem to have existed at the middle of the Mesozoic age—not until towards its close (in the Cretaceous period). At all events, we have no fossil remains of indubitable Placentals from that period.
The existing Marsupials, of which the plant-eating kangaroo and the carnivorous opossum (Fig. 272) are the best known, differ a good deal in structure, shape, and size, and correspond in many respects to the various orders of Placentals. Most of them live in Australia, and a small part of the Australian and East Malayan islands. There is now not a single living Marsupial on the mainland of Europe, Asia, or Africa. It was very different during the Mesozoic and even during the Cenozoic age. The sedimentary deposits of these periods contain a great number and variety of marsupial remains, sometimes of a colossal size, in various parts of the earth, and even in Europe. We may infer from this that the existing Marsupials are the remnant of an extensive earlier group that was distributed all over the earth. It had to give way in the struggle for life to the more powerful Placentals during the Tertiary period. The survivors of the group were able to keep alive in Australia and South America because the one was completely separated from the other parts of the earth during the whole of the Tertiary period, and the other during the greater part of it.
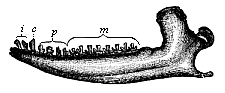 Fig. 271—Lower jaw of a Promammal (Dryolestes priscus), from the Jurassic of the Felsen strata. (From Marsh.)
Fig. 271—Lower jaw of a Promammal (Dryolestes priscus), from the Jurassic of the Felsen strata. (From Marsh.) |
From the comparative anatomy and ontogeny of the existing Marsupials we may draw very interesting conclusions as to their intermediate position between the earlier Monotremes and the later Placentals. The defective development of the brain (especially the cerebrum), the possession of marsupial bones, and the simple construction of the allantois (without any placenta as yet) were inherited by the Marsupials, with many other features, from the Monotremes, and preserved. On the other hand, they have lost the independent bone (caracoideum) at the shoulder-blade. But we have a more important advance in the disappearance of the cloaca; the rectum and anus are separated by a partition from the uro-genital opening (sinus urogenitalis). Moreover, all the Marsupials have teats on the mammary glands, at which the new-born animal sucks. The teats pass into the cavity of a pouch or pocket on the ventral side of the mother, and this is supported by a couple of marsupial bones. The young are born in a very imperfect condition, and carried by the mother for some time longer in her pouch, until they are fully developed (Fig. 272). In the giant kangaroo, which is as tall as a man, the embryo only develops for a month in the uterus, is then born in a very imperfect state, and finishes its growth in the mother’s pouch (marsupium); it remains in this about nine months, and at first hangs continually on to the teat of the mammary gland.
From these and other characteristics (especially the peculiar construction of the internal and external sexual organs in male and female) it is clear that we must conceive the whole sub-class of the Marsupials as one stem group, which has been developed from the Promammalia. From one branch of these Marsupials (possibly from more than one) the stem-forms of the higher Mammals, the Placentals, were afterwards evolved. Of the existing forms of the Marsupials,
which have undergone various modifications through adaptation to different environments, the family of the opossums (Didelphida or Pedimana) seems to be the oldest and nearest to the common stem-form of the whole class. To this family belong the crab-eating opossum of Brazil (Fig. 272) and the opossum of Virginia, on the embryology of which Selenka has given us a valuable work (cf. Figs. 63–67 and 131–135). These Didelphida climb trees like the apes, grasping the branches with their hand-shaped hind feet. We may conclude from this that the stem-forms of the Primates, which we must regard as the earliest Lemurs, were evolved directly from the opossum. We must not forget, however, that the conversion of the five-toed foot into a prehensile hand is polyphyletic. By the same adaptation to climbing trees the habit of grasping their branches with the feet has in many different cases brought about that opposition of the thumb or great toe to the other toes which makes the hand prehensile. We see this in the climbing lizards (chameleon), the birds, and the tree-dwelling mammals of various orders.
 Fig. 272—The crab-eating Opossum (Philander cancrivorus). The female has three young in the pouch. (From Brehm.)
Fig. 272—The crab-eating Opossum (Philander cancrivorus). The female has three young in the pouch. (From Brehm.) |
Some zoologists have lately advanced the opposite opinion, that the Marsupials represent a completely independent
sub-class of the Mammals, with no direct relation to the Placentals, and developing independently of them from the Monotremes. But this opinion is untenable if we examine carefully the whole organisation of the three sub-classes, and do not lay the chief stress on incidental features and secondary adaptations (such as the formation of the marsupium). It is then clear that the Marsupials—viviparous Mammals without placenta—are a necessary transition from the oviparous Monotremes to the higher Placentals with chorion-villi. In this sense the Marsupial class certainly contains some of man’s ancestors.
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XXI
Chapter XXIII
Figs. 1–209
Figs. 210–408